
 United States
United States
 Germany
Germany
 Japan
Japan
 United Kingdom
United Kingdom
 China
China
Melanoma Antigen (MAGE) Family - More than a Cancer Cell Marker
The melanoma antigen (MAGE) protein family, containing a ~180-amino-acid MAGE homology domain (MHD), are conserved in eukaryotes and have ~40 members in human and mouse. MAGEA1, the founder member of this family, was first identified as a cancer cell marker in melanoma, and soon after, it was found having restricted expression in male gonad, a mammalian body sites with immune privilege. Due to the antigenic potential, male germline expression, and aberrantly re-activated in cancer cells, two third of MAGE proteins have been categorizes as cancer-testis antigens (CTA).
OriGene offers a range of tools for analyzing MAGE Protein Family. Click below for details
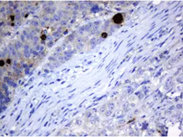
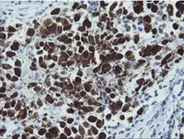
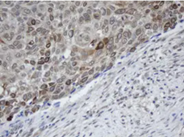
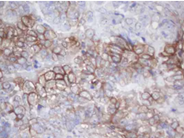
MAGE Proteins: Normal function and Implication in Diseases
The physiological functions of MAGE proteins are diverse. For example, MAGEAs enhance the resistance to the genotoxic stress during spermatogenesis. MAGEB2 maintains thermo-sensitive spermatogenesis by suppressing stress granule formation. MAGED1 regulates neuronal apoptosis during neurogenesis and plays essential roles in various brain functions, such as depression, memory, behavior, and circadian rhythm. MAGED2, mutations of which cause X-linked polyhydramnios with prematurity and a transient form of antenatal Bartter's syndrome, regulates expression and function of Na-K-Cl cotransporter (NKCC2) and Na-Cl cotransporter (NCC). Children with MAGEG1 mutations suffered from lung disease immunodeficiency and chromosome breakage syndrome. MAGEL2 is implicated in various human genetic disorders, e.g. Prader-Willi syndrome and Schaaf-Yang syndrome.
MAGE Proteins in Cancers
MAGE proteins have widespread expression in different cancers and are usually poor prognostic markers for cancer patients. As convincing cancer cell markers, MAGE proteins (e.g. MAGEA1, A3, A4, A6, A10, A12, C2) have been immuno-targeted in many clinical trials for cancer. However, myriad evidence indicates that MAGE proteins also play an active role in oncogenicity, tumor progression, and cancer cell survival. For example, MAGEA3 can suppresses cancer cell apoptosis through directly interacting with TP53. MAGEA3 can also complex with TRIM28 to ubiquitinate and degrade TP53 or AMPKα, a master sensor/regulator of cell energy homeostasis, which is commonly suppressed in early-stage cancers. MAGEA11 complexed with HUWE1 performs ubiquitin E3 ligase activity to degrade PCF11, a mRNA 3' end processing complex protein, resulting in 3' UTR shortening, a common feature of most cancer cells. MAGEF1-NSMCE1 E3 ubiquitin ligases ubiquitinate and degrade MMS19, an essential protein in cytosolic iron-sulfur (Fe-S) cluster assembly (CIA) pathway, leading to defects in DNA repair and increasing mutation burden in cancer cells. MAGE proteins should be considered as new therapeutic targets for cancers.
MAGEA Antibody Panel
OriGene offers a range of tools for analyzing MAGE Protein Family. Click the boxes for details
MAGE in Clinical Trial
A1 A3 A4 A6 A10 A12 C2MAGE - Ubiquitin E3 Ligase
TRIM28 RAD18 HUWE1 SKIP2 LNX1 RBX1 PJA1 NSMCE1 TRIM27 RNF41MAGE Family
A1 A2 A3 A4 A5 A6 A8 A9 A10 A11 A12 B1 B2 B3 B4 B5 B6 B10 B16 B17 B18 C1 C2 C3 D1 D2 D3 D4 E1 E2 F1 G1 H1 L2 NDN| Gene | CTA* | Function | Associated proteins | Activity | Target | Pathway | Reference (PubMed ID) |
|---|---|---|---|---|---|---|---|
| MAGEA2 | x | transcription | TP53, HDAC3 | transcription repressor | CDKN1A | cell cycle regulation | 16847267 |
| transcription | chromatin binding blocker | TP53 | tumor suppression | 21056992 | |||
| protein degradation | TRIM28 | E3 ubiquitin ligase | TP53 | tumor suppression | 20864041 | ||
| MAGEA3/6 | x | protein degradation | TRIM28 | E3 ubiquitin ligase | AMPK alpha (PRKAA1) | cell metabolism | 25679763 |
| protein degradation | TRIM28 | E3 ubiquitin ligase | ZNF382 | cell cycle regulation | 21876767 | ||
| protein degradation | TRIM28 | E3 ubiquitin ligase | FBP1 | cell metabolism | 28394358 | ||
| unknown | negetive regulator | BIRC5 | apoptosis | 34166362 | |||
| MAGEA4 | x | protein degradation | RAD18 | E3 ubiquitin ligase | PCNA | Trans-lesion synthesis (TLS) | 27377895 |
| MAGEA11 | x | protein degradation | HUWE1 | E3 ubiquitin ligase | PCF11 | mRNA alternative polyadenylation (APA) | 31980388 |
| Transcription | AR, EP300, NCOA2 | transcription activator | KLK3 | androgen response | 20448036 | ||
| Transcription | PGR (B isoform) | transcription activator | FKBP5 | progesteron response | 22891251 | ||
| protein degradation/ stabilization | SKP2 (SCF complex) | E3 ubiquitin ligase | CCNA2, E2F1, RBL1 | cell cycle regulation | 27720894 | ||
| MAGEB2 | x | Translation | translation repressor (DDX5 antagonist) | G3BP1 | stress granule formation | 32692974 | |
| Transcription | HDAC1 | transcription activator | E2F1 | cell cycle regulation | 26468294 | ||
| Transcription | rRNA transcription pre-initiation complex | UBTF | ribosome biogenesis | 33741433 | |||
| MAGEB18 | x | protein degradation | LNX1 | E3 ubiquitin ligase | unknown | 20864041 | |
| MAGEC2 | x | protein degradation | TRIM28 | E3 ubiquitin ligase | AMPK alpha (PRKAA1) | cell metabolism | 25679763 |
| protein degradation | TRIM28 | E3 ubiquitin ligase | TP53 | tumor suppression | 20864041 | ||
| protein degradation | TRIM28 | E3 ubiquitin ligase | ZNF382 | cell cycle regulation | 21876767 | ||
| protein degradation | ubiquitination | ZMYND11 | tumor suppression | 24866244 | |||
| protein stabilization | polyubiquitination inhibition | STAT3 | cancer metastasis | 27775077 | |||
| protein stabilization | RBX1, Skp1-Cullin1-F box protein (SCF) | E3 ubiquitin ligase inhibition | CCNE1 | cell cycle regulation | 26540345 | ||
| protein degradation | TRIM28 | E3 ubiquitin ligase | FBP1 | cell metabolism | 28394358 | ||
| MAGED1 | Signaling | NGFR | signaling adaptor | JNK | neuronal apoptosis | 12376548 | |
| Signaling | UNC5A | signaling adaptor | neuronal apoptosis | 12598531 | |||
| protein degradation | pro-apoptotic protein | XIAP | neuronal apoptosis | 11546791 | |||
| protein degradation | pro-apoptotic protein | AATF | neuronal apoptosis | 17488777 | |||
| Signaling | TAK1, TAB1, XIAP | signaling complex | MAPK14 | apoptosis | 16107717 | ||
| Transcription | transcription coactivator | DLX5 | cell differentiation | 11084035 | |||
| protein degradation | PJA1 | E3 ubiquitin ligase | unkown | 20864041 | |||
| transcription | NDN | transcription factor modulator | MSX1 | cell differentiation | 15272023 | ||
| protein degradation | ubiquitination | SLC6A4 | neurotransmitter serotonin retrieval | 22457503 | |||
| transcription | RORA | transcription coactivator | ARNTL | circadian rhythm | 20300063 | ||
| transcription | CREB1 | transcription coactivator | BDNF, ATF4, NRXN1, GRIN2B, EGR1 | synaptic plasticity | 24700102 | ||
| MAGED2 | unkown | NCL | unkown | cell cycle: G1/S transition | 29371029 | ||
| transcription | TP53 | transcription factor modulator | apoptosis | 17912449 | |||
| protein stablization | GNAS, DNAJB1 | unkown | SLC12A1, SLC12A3 | Bartter's syndrome | 27120771 | ||
| MAGEF1 | protein degradation | NSMCE1 | E3 ubiquitin ligase | MMS19 | cytosolic iron-sulfur (Fe-S) cluster assembly (CIA) pathway | 29225034 | |
| MAGEG1 (NDNL2, NSMCE3) | protein complex | SMC5/6 complexes | DNA replication fork stabilizer | DNA replication and repair | 17005570 | ||
| protein degradation | NSMCE1 | E3 ubiquitin ligase | unknown | 20864041 | |||
| MAGEH1 | unkown | GADD45G | caspases activation | apoptosis | 34788311 | ||
| MAGEL2 | protein degradation | E3 ubiquitin ligase | CRY1 | circadian rhythm | 32315313 | ||
| mRNA processing | mRNA m6A methylation | YTHDF2 | RNA metabolism | 34265304 | |||
| signaling | TRIM27, USP7 | E3 ubiquitin ligase | WASHC1 | endosomal protein recycling | 26365382 | ||
| signaling | NDN, RNF41, USP8 | E3 ubiquitin ligase | LEPR | endosomal protein recycling | 28973533 | ||
| NDN | transcription | SIRT1 | deacetylase | TP53 | neuronal apoptosis | 18753379 |
*Cancer Testis Antigen
References
- Florke Gee et al. 2020 Nov 20;295(47):16121-16155. doi: 10.1074/jbc.REV120.008029
- Hasegawa and Yoshikawa. 2008 Aug 27;28(35):8772-84. doi: 10.1523/JNEUROSCI.3052-08.2008
- Shin et al. 2021 Nov 17;16(11):e0260135. doi: 10.1371/journal.pone.0260135
- Sanderson et al. 2021 Aug;297(2):100959. doi: 10.1016/j.jbc.2021.100959
- Ladelfa et al. 2021 Jun;1868(7):119015. doi: 10.1016/j.bbamcr.2021.119015
- Craig et al. 2021 Jun 24;17(6):e1009589. doi: 10.1371/journal.pgen.1009589
